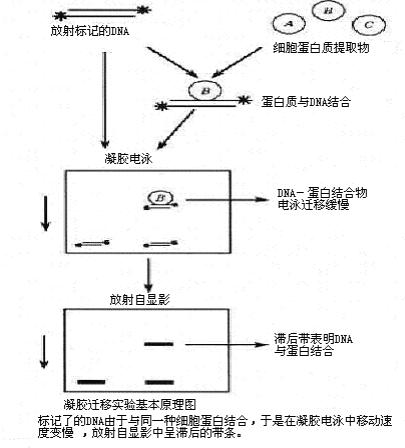
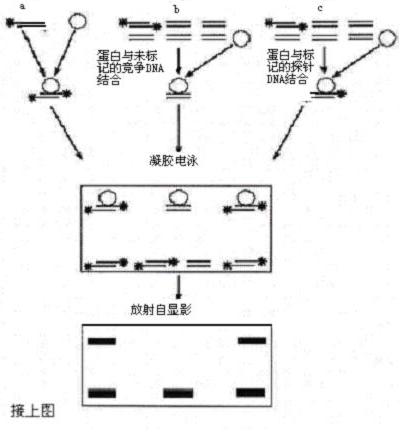
凝胶迁移或电泳迁移率实验(EMSA)是一种用于研究核酸(DNA或RNA)与蛋白质相互作用的经典分子生物学技术。该方法通过非变性聚丙烯酰胺凝胶电泳,将蛋白质与核酸形成的复合物与游离核酸分离,进而分析结合特异性和结合强度。EMSA广泛用于转录因子、RNA结合蛋白等调控因子的功能研究。
实验原理与流程:
EMSA通常将纯化蛋白或细胞抽提液与放射性或荧光标记的DNA/RNA探针孵育,形成复合物后进行非变性凝胶电泳。结合后的核酸迁移速度减慢,形成特异性迁移带。通过竞争实验(特异性与非特异性竞争核酸)可进一步验证结合的特异性。
所需试剂与系统:
实验需准备结合蛋白(纯化、部分纯化或抽提液)、同位素或荧光标记的DNA/RNA探针、结合缓冲液(含盐、缓冲体系、还原剂、甘油、非特异竞争DNA等)。常用标记方法包括g-32P标记DNA、体外转录标记RNA。Promega等公司提供EMSA专用试剂盒,包括正对照蛋白、寡核苷酸探针、结合缓冲液等。
优化因素:
EMSA实验需优化蛋白与探针浓度、缓冲液配方、pH、凝胶浓度、电泳条件、保温时间与温度、载体蛋白及辅助因子。常用缓冲液含4-10%甘油、1mM MgCl2、0.5mM EDTA、0.5mM DTT、50mM NaCl、10mM Tris-HCl(pH7.5)等。
探针设计与选择:
探针长度一般小于300bp,常用25bp左右的寡核苷酸以便区分结合位点。长片段可用于定位蛋白结合区域,随后可用DNase I印迹分析结合位点。
常见转录因子与复合物:
EMSA可检测AP1、AP2、CREB、NF-κB、Oct1、Sp1、TFIID/TFIIB等转录因子与其同源结合序列形成的复合物。每种因子具有特定的结合序列和复合物迁移特征。实验条件需根据因子特性调整。
竞争DNA与poly(dI:dC):
poly(dI:dC)由肌苷和胞嘧啶组成,用于抑制蛋白对探针的非特异结合,避免假复合物。特异性竞争DNA与非特异性竞争DNA用于验证结合特异性。
凝胶条件与分离:
非变性聚丙烯酰胺凝胶(6%)常用于分离复合物与游离探针。凝胶浓度、pH、电泳电压需根据蛋白与探针特性优化。低温操作可防止复合物解离。
复合物鉴定:
可通过超迁移实验(抗体干扰)、UV交联、Western印迹等方法确定复合物中蛋白的身份。竞争实验和突变分析可进一步验证结合序列。
参考文献:
Briggs MR et al. Science 234, 47 (1986); Lee W et al. Cell 49, 741 (1986); Williams T et al. Genes Dev 2, 1557 (1989); Sen R and Baltimore D. Cell 46, 705 (1986); Parlsow TG et al. Proc Natl Acad Sci USA 81, 2650 (1984); Montminy MR et al. Proc Natl Acad Sci USA 83, 6682 (1986); Angel P et al. Cell 49, 729 (1987); Chiu R et al. Cell 54, 541 (1988); Rauscher FJ et al. Cell 52,471 (1988); Imagawa M et al. Cell 51, 251 (1987); Berkowitz LA and Gilman MZ. Proc Natl Acad Sci USA 87, 5258 (1990); Baeuerle PA. Biochim Biophys Acta 1071, 63 (1991); Duckett CS et al. Mol Cell Biol 13, 1315 (1993); Urban MB et al. EMBO J 10(7), 1817 (1991); Dynan WS and Tjian R. Cell 35, 79 (1983); Peterson MG et al. Science 248, 1652 (1990); Coleman RA et al. J Biol Chem 270, 13842 (1995); Beckler G and Hurst R. Promega Notes 43,24 (1993); DiDonato JA and Karin M. Promega Notes 42, 18 (1993); Demczuk S et al. Proc Natl Acad Sci USA90,2574 (1993)。
使用Promega公司EMSA产品的参考文献:Xu J and Clark RAF. J Cell Biol 136, 473 (1997); Dbaibo GS et al. J Exp Med 185, 481 (1997); Pan ZK et al. J Clin Invest 98, 2042 (1996); Kochekova M et al. J Clin Invest 99, 3000 (1997); Avantaggiati ML et al. Cell 89, 1175 (1997); Schmedite JF et al. J Biol Chem 272, 601 (1997); Lee BS et al. J Biol Chem 272, 174 (1997); Ohlsson BG et al. J Clin Invest 98, 78 (1997); In KH et al. J Clin Invest 99, 1130 (1997)。