|
4.在体外实验中,将线粒体分成各种复合物,检测其各自催化的反应,再将其重组,检测其催化能力。 美国格林(Green)等实验室成功地将呼吸链分离成具有催化活性的四种复合物以及CoQ和Cytc.检测各个复合物的功能发现: 可以看出CoQ在复合物Ⅰ与Ⅲ,Ⅱ与Ⅲ之间传递还原当量,Cyt c在复合物Ⅲ与Ⅳ之间传递还原当量。他们又将这四种复合物1:1:1:1的比例混合,加上CoQ和Cyt c重组,基本上恢复了线粒体原有的催化能力。 借助上述实验方法,呼吸链各组分的排列顺序已基本明确,但仍有些不一致的看法,其中以CoQ至细胞色素C这一部分研究得还很不清楚,对于Fe-S和CoQ的定位和数量也有争议。 (二)氧化呼吸链 1.NADH氧化呼吸链 人体内大多数脱氢酶都以NAD+作辅酶,在脱氢酶催化下底物SH2脱下的氢交给NAD+生成NADH+H+,在NADH脱氢酶作用下,NADH+H+将两个氢原子传递给FMN生成FMNH2,再将氢传递至CoQ生成CoQH2,此时两个氢原子解离成2H++2e,2H+游离于介质中,2e经Cyt b、c1、c、aa3传递,最后将2e传递给1/2O2,生成O2-,O2与介质中游离的2H+结合生成水,综合上述传递过程可用图6-6表示。
图6-6 NADH氧化呼吸链? SH2:作用物;(Fe-S):铁硫中心;Cyt:细胞色素 2.琥珀酸氧化呼吸链 琥珀酸在琥珀酸脱氢酶作用下脱氢生成延胡索酸,FAD接受两个氢原子生成FADH2,然后再将氢传递给CoQ,生成CoQH2,此后的传递和NADH氧化呼吸链相同,整个传递过程可用图6-7表示。
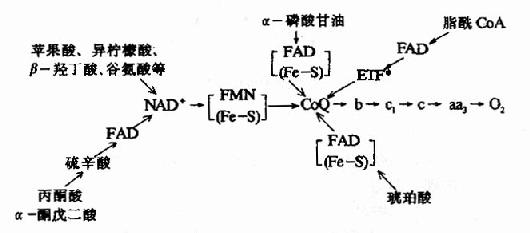
图6-7 琥珀酸氧化呼吸链? (Fe-S):铁硫中心:b:琥珀酸脱氢酶复合体的细胞色素 3.线粒体氧化呼吸链总结 线粒体中物质代谢会生成大量的NADH+H+和FADH2-它们可来自丙酮酸氧化脱羧、三羧酸循环、脂肪酸的β-氧化和L-谷氨酸的氧化脱氨等反应,现将某些重要底物氧化时的呼吸链总结于图6-8。
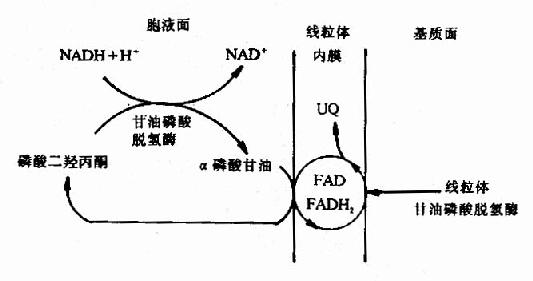
图6-8 线粒体中某些底物氧化时的呼吸链 ?*ETF:电子传递黄素蛋白,辅基为FAD 三、胞浆中NADH的转移 体内很多物质氧化分解产生NADH,反应发生在线粒体内,则产生的NADH可直接通过呼吸链进行氧化磷酸化,但亦有不少反应是在线粒体外进行的,如3-磷酸甘油醛脱氢反应,乳酸脱氢反应及氨基酸联合脱氨基反应等等。由于所产生的NADH存在于线粒体外,而真核细胞中,NADH不能自由通过线粒体内膜,因此,必须借助某些能自由通过线粒体内膜的物质才能被转入线粒体,这就是所谓穿梭机制,体内主要有两种穿梭机制。 1.α磷酸甘油穿梭(glycerol?α-phosphate shuttle) 该穿梭机制主要在脑及骨骼肌中,它是借助于α-磷酸甘油与磷酸二羟丙酮之间的氧化还原转移还原当量,使线粒体外来自NADH的还原当量进入线粒体的呼吸链氧化,具体过程如图6-9。
图6-9 α磷酸甘油穿 当胞液中NADH浓度升高时,胞液中的磷酸二羟丙酮首先被NADH还原成α磷酸甘油(3-磷酸甘油),反应由甘油磷酸脱氢酶(辅酶为NAD+)催化,生成的α磷酸甘油可再经位于线粒体内膜近外侧部的甘油磷酸脱氢酶催化氧化生成磷酸二羟丙酮。线粒体与胞液中的甘油磷酸脱氢酶为同工酶,两者不同在于线粒体内的酶是以FAD为辅基的脱氢酶,而不是NADH+,FAD所接受的质子、电子可直接经泛醌、复合体Ⅲ、Ⅳ传递到氧,这样线粒体外的还原当量就被转运到线粒体氧化了,但通过这种穿梭机制果只能生成2分子ATP而不是3分子ATP。 2.苹果酸,天冬氨酸穿梭(malate aspartate shuttle): 这种穿梭机制主要在肝、肾、心中发挥作用,其穿梭机制比较复杂,不仅需借助苹果酸、草酸乙酸的氧化还原,而且还要借助α酮酸与氨基酸之间的转换,才能使胞液中来的NADH的还原当量转移进入线粒体氧化,具体过程如图6-10。
图6-10 苹果酸天冬氨酸穿梭? GOT:谷草转氨酸;MDH:苹果酸脱氢酶 当胞液中NADH浓度升高时,首先还原草酰乙酸成为苹果酸,此反应由苹果酸脱氢酶催化,胞液中增多的苹果酸可通过内膜上的二羧酸载体系统与线粒体内的α酮戊二酸交换;进入线粒体的苹果酸,经苹果酸脱氢酶催化又氧化生成草酰乙酸并释出NADH,还原当量从复合体I进入呼吸链经CoQ、复合体Ⅲ、Ⅳ传递,最image/005061360后给氧,所以仍可产生3分子ATP,与在线粒体内产生的NADH氧化相同。与此同时线粒体内的α酮戊二酸由于与苹果酸交换而减少,需要补充,于是在转氨酶作用下由谷氨酸与草酰乙酸进行转氨基反应,生成α酮戊二酸和天冬氨酸,天冬氨酸借线粒体膜上的谷氨酸天冬氨酸载体转移系统与胞液的谷氨酸交换,从而补充了线粒体内谷氨酸由于转氨基作用而造成的损失,进入胞液的天冬氨酸再与胞液中α酮戊二酸进行转氨基,重新又产生草酰乙酸以补充最初的消耗,从而完成整个穿梭过程。? (责任编辑:泉水) |
第二节 呼吸链(2)
时间:2006-06-20 10:47来源:大众医药网 作者:admin 点击:
924次
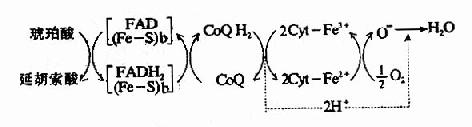


顶一下
(5)
100%
踩一下
(0)
0%
------分隔线----------------------------
- 发表评论
-
- 最新评论 进入详细评论页>>
