|
嗜盐菌(halobacterium haloblum)是一种能在高浓度盐溶液中生长的细菌,该菌中有一种结合蛋白质,称为菌紫质(bacteriorhodopsin),菌紫质能将光能转换成化学能。有人使用嗜盐菌作实验,在无O2的情况下用光照射嗜盐菌,尽管无氧化作用,菌体内仍维持一定的ATP浓度,若加入解偶联剂或加入磷酸化抑制剂DCC,则菌体内ATP浓度降低;而加入呼吸抑制剂抑制电子传递,即不影响ATP合成,ATP浓度不变,这说明电子传递和H+运动是可以分开加以研究的,嗜盐菌为研究化学渗透学说的H+运动提供了一个理想的模型。于是,有人分离嗜盐菌的菌紫质,并将其重组在人工脂质体中,然后用光照射,可测得跨膜电位为?120mV(内负外正),同时膜外侧H+浓度增高,膜内外△pH约为1.8单位,可以算出总的质子移动力约为△P=-120mV-59×1.8mV=226mV,若再将牛心线粒体内膜重组在此脂质体中,光照后可使ADP+Pi生成ATP,这说明质子跨膜梯度可以经过线粒体内膜的三分子体将H+跨膜梯度中储存的能量转变为ATP分子中的化学能。 五、氧化磷酸化抑制剂 氧化磷酸化抑制剂可分为三类,即呼吸抑制剂、磷酸化抑制剂和解偶联剂。 (一)呼吸抑制剂 这类抑制剂抑制呼吸链的电子传递,也就是抑制氧化,氧化是磷酸化的基础,抑制了氧化也就抑制了磷酸化。呼吸链某一特定部位被抑制后,其底物一侧均为还原状态,其氧一侧均为氧化态,这很容易用分光光度法(双波长分光光度计)检定,重要的呼吸抑制剂有以下几种。 鱼藤酮(rotenone)系从植物中分离到的呼吸抑制剂,专一抑制NADH→CoQ的电子传递。 抗霉素A(actinomycin A)由霉菌中分离得到,专一抑制CoQ→Cyt c的电子传递。 CN、CO、NaN3和H2S均抑制细胞色素氧化酶。 (二)磷酸化抑制剂 这类抑制剂抑制ATP的合成,抑制了磷酸化也一定会抑制氧化。 寡霉素(oligomycin)可与F0的OSCP结合,阻塞氢离子通道,从而抑制ATP合成。 二环己基碳二亚胺(dicyclohexyl carbodiimide,DCC)可与F0的DCC结合蛋白结合,阻断H+通道,抑制ATP合成。栎皮酮(quercetin)直接抑制参与ATP合成的ATP酶。 (三)解偶联剂(uncoupler) 解偶联剂使氧化和磷酸化脱偶联,氧化仍可以进行,而磷酸化不能进行,解偶联剂作用的本质是增大线粒体内膜对H+的通透性,消除H+的跨膜梯度,因而无ATP生成,解偶联剂只影响氧化磷酸化而不干扰底物水平磷酸化,解偶联剂的作用使氧化释放出来的能量全部以热的形式散发。动物棕色脂肪组织线粒体中有独特的解偶联蛋白,使氧化磷酸化处于解偶联状态,这对于维持动物的体温十分重要。 常用的解偶联剂有2,4-二硝基酚(dinitrophenol,DNP),羰基-氰-对-三氟甲氧基苯肼(FCCP),双香豆素(dicoumarin)等,过量的阿斯匹林也使氧化磷酸化部分解偶联,从而使体温升高。 过量的甲状腺素也有解偶联作用,甲状腺素诱导细胞膜上Na+-K+-ATP酶的合成,此酶催化ATP分解,释放的能量将细胞内的Na+泵到细胞外,而K+进入细胞,Na+-K+-ATP酶的转换率为100个分子ATP/秒,酶分子数增多,单位时间内分解的ATP增多,生成的ADP又可促进磷酸化过程。甲亢病人表现为多食、无力、喜冷怕热,基础代谢率(BMR)增高,因此也有人将甲状腺素看作是调节氧化磷酸化的重要激素。 六、氧化磷酸化的调节 机体的氧化磷酸化主要受细胞对能量需求的调节 (一)ATP/ADP值对氧化磷酸化的直接影响 线粒体内膜中有腺苷酸转位酶,催化线粒体内ATP与线粒体外ADP的交换,ATP分子解离后带有4个负电荷,而ADP分子解离后带有3个负电荷,由于线粒体内膜内外有跨膜电位(△ψ),内膜外侧带正电,内膜内侧带负电,所以ATP出线粒体的速度比进线粒体速度快,而ADP进线粒体速度比出线粒体速度快。Pi进入线粒体也由磷酸转位酶催化,磷酸转位酶催化OH?与Pi交换,磷酸?二羧酸转位酶催化Pi2-与二羧酸(如苹果酸)交换。 当线粒体中有充足的氧和底物供应时,氧化磷酸化就会不断进行,直至ADP+Pi全部合成ATP,此时呼吸降到最低速度,若加入ADP,耗氧量会突然增高,这说明ADP控制着氧化磷酸化的速度,人们将ADP的这种作用称为呼吸受体控制。 机体消耗能量增多时,ATP分解生成ADP,ATP出线粒体增多,ADP进线粒体增多,线粒体内ATP/ADP值降低,使氧化磷酸化速度加快,ADP+Pi接受能量生成ATP。机体消耗能量少时,线粒体内ATP/ADP值增高,线粒体内ADP浓度减低就会使氧化磷酸化速度减慢。 (二)ATP/ADP值的间接影响 ATP/ADP值增高时,使氧化磷酸化速度减慢,结果NADH氧化速度减慢,NADH浓度增高,从而抑制了丙酮酸脱氢酶系、异柠檬酸脱氢酶、α-酮戊二酸脱氢酶系和柠檬酸合成酶活性,使糖的氧化分解和TCA循环的速度减慢。 (三)ATP/ADP值对关键酶的直接影响 ATP/ADP值增高会抑制体内的许多关键酶,如变构抑制磷酸果糖激酶、丙酮酸激酶和异柠檬酸脱氢酶,还能抑制丙酮酸脱羧酶、α-酮戊二酸脱氢酶系,通过直接反馈作用抑制糖的分解和TCA循环。 七、高能磷酸化合物的储存和利用 无论是底物水平磷酸化还是氧化磷酸化,释放的能量除一部分以热的形式散失于周围环境中之外,其余部分多直接生成ATP,以高能磷酸键的形式存在。同时,ATP也是生命活动利用能量的主要直接供给形式。 (一)高能化合物 人体存在多种高能化合物,但这些高能化合物的能量并不相同。 体外实验中,在pH7.0,25℃条件下,每克分子ATP水解生成ADP+Pi时释放的能量为7.1千卡或30.4千焦耳,在体内,pH7.4,37℃,ATP、ADP+Pi、Mg2+均处于细胞内生理浓度的情况下,每克分子ATP水解生成ADP+Pi时释放的能量为33.5-50千焦耳或8-12千卡(表6-4)。 ?表6-4 几种常见高能化合物水解时释放的能量
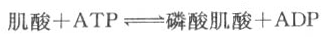
卫生学规定,中度体力劳动者每日每kg体重需供给能量34-40千卡,若一成人重70kg,从事中度体力劳动,则每日应供应含能量2450千卡的食物,其中40%的能量转变成化学能储存于ATP分子的高能键中,这一部分能量应为2450×0.4=980.0千卡,按每克分子ATP水解生成ADP+Pi释放7.3千卡能量计算,应当合成:980÷7.3=134.3克分子ATP,ATP的分子量为507.22,所以134.3克分子ATP重达68.12kg,这足以表明ATP在体内的代谢十分旺盛。 ATP在能量代谢中之所以重要,就是因为ATP水解时的标准自由能变化位于多种物质水解时标准自由能变化的中间,它能从具有更高能量的化合物接受高能磷酸键,如接受PEP、1,3-二磷酸甘油、磷酸肌酸分子中的~Pi生成ATP,ATP也能将~Pi转移给水解时标准自由能变化较小的化合物,如转移给葡萄糖生成G-6-P。 (二)ATP能量的转移 ATP是细胞内的主要磷酸载体,ATP作为细胞的主要供能物质参与体内的许多代谢反应,还有一些反应需要UTP或CTP作供能物质,如UTP参与糖元合成和糖醛酸代谢,GTP参与糖异生和蛋白质合成,CTP参与磷脂合成过程,核酸合成中需要ATP、CTP、UTP和GTP作原料合成RNA,或以dATP、dCTP、dGTP和dTTP作原料合成DNA。 作为供能物质所需要的UTP、CTP和GTP可经下述反应再生: UDP+ATP→UTP+ADP GDP+ATP→GTP+ADP CDP+ATP→CTP+ADP dNTP由dNDP的生成过程也需要ATP供能: dNDP+ATP→dNTP+ADP (三)磷酸肌酸 ATP是细胞内主要的磷酸载体或能量传递体,人体储存能量的方式不是ATP而是磷酸肌酸。肌酸主要存在于肌肉组织中,骨骼肌中含量多于平滑肌,脑组织中含量也较多,肝、肾等其它组织中含量很少。 磷酸肌酸的生成反应如下:
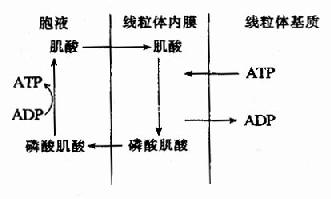
肌细胞线粒体内膜和胞液中均有催化该反应的肌酸激酶,它们是同工酶。线粒体内膜的肌酸激酶主要催化正向反应,生成的ADP可促进氧化磷酸化,生成的磷酸肌酸逸出线粒体进入胞液,磷酸肌酸所含的能量不能直接利用;胞液中的肌酸激酶主要催化逆向反应,生成的ATP可补充肌肉收缩时的能量消耗,而肌酸又回到线粒体用于磷酸肌酸的合成,此过程可用图6-15表示。
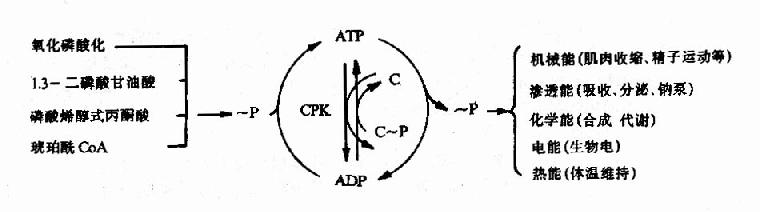
图6-15 磷酸肌酸的生成与利用 肌肉中磷酸肌酸的浓度为ATP浓度的5倍,可储存肌肉几分钟收缩所急需的化学能,可见肌酸的分布与组织耗能有密切关系。 ATP的生成、储存和利用可用下图表示(图6-16)
图6-16 ATP的生成、储存和利用总结示意图? CPK:肌酸磷酸激酶 (责任编辑:泉水) |
第三节 ATP的生成、储存和利用(2)
时间:2006-06-20 10:47来源:大众医药网 作者:admin
顶一下
(4)
100%
踩一下
(0)
0%
------分隔线----------------------------
- 上一篇:第七章 氨基酸代谢- -第一节 氨基酸的一般代谢
- 下一篇:第二节 呼吸链
- 发表评论
-
- 最新评论 进入详细评论页>>
